Application Note: High-Resolution Imaging of DNA Nanoarchitectures Using AFM
Decoding the human genome was one of the major scientific aims of the 20th century, with the idea that understanding the code of life will lead to an understanding of the human organism and specific problems like genetic diseases. In the past decades, atomic force microscopy (AFM) has become standard in the high-resolution structural analysis of samples ranging from single molecules to complex macromolecular systems, such as DNA..
In this application note, Bruker's NanoWizard ULTRA Speed* AFM was used to study DNA-based structures, such as the general structure of a DNA double-helix, tile-based DNA arrays, and DNA origami.
*The latest generation system in this product line is the NanoWizard ULTRA Speed 3
Readers can expect to learn about:
- DNA nanostructures, including tile-based DNA arrays and folding-based DNA origami, and how they are designed and self-assembled through complementary overhangs and scaffold folding;
- Comprehensive application examples showcasing the high-resolution imaging capabilities of Bruker’s NanoWizard ULTRA Speed AFM, revealing details of major and minor grooves, helix pitch, and base pairing; and
- The potential of the NanoWizard ULTRA Speed as a powerful tool for real-time imaging and capturing dynamic processes in delicate biological samples.
KEYWORDS: Atomic Force Microscopy; Fast Scanning AFM; High-Resolution Imaging; DNA origami; Life Sciences
Introduction
All the instructions used for the development and functioning of living organisms are stored in the DNA macromolecule. Decoding the human genome was one of the major scientific aims of the 20th century. The fascinating idea is to understand the code of life and get a better understanding of the human organism and specific problems like human genetic diseases and the possibilities of avoiding and curing such diseases. Other questions also follow on from these discoveries. How is it possible that the complexity of a human brain is stored in such a simply structured DNA molecule? Is it possible to use the genetic code to produce patient-specific drugs?
LEARN MORE:
The structure of linear double-stranded DNA
To decode the DNA sequence the nucleic acid was intensively investigated by many research groups around the world over the last 150 years. The double-helix structure of B-DNA was discovered by Watson and Crick, for which they received a Nobel Prize in 1962 with Wilkins.
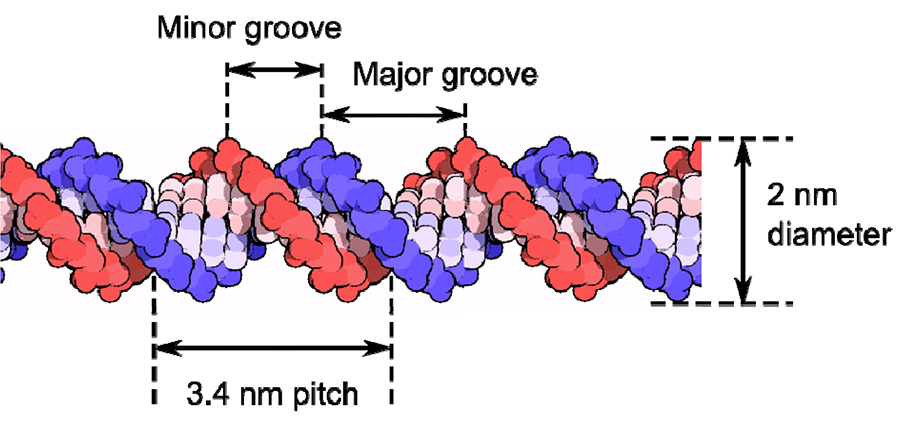
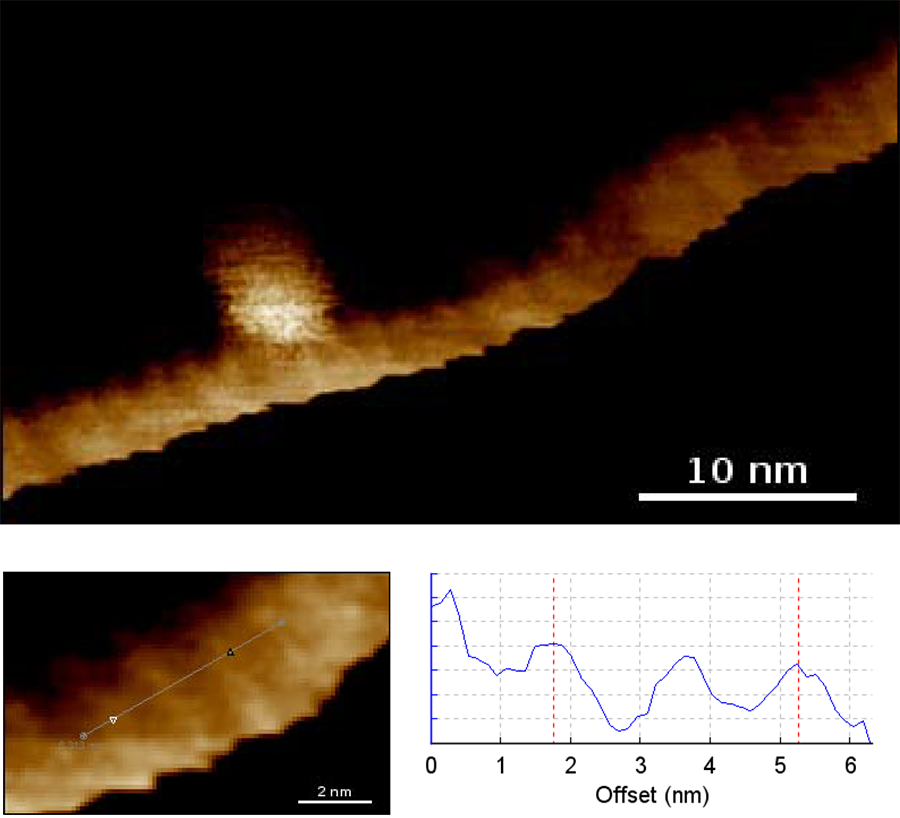
The structure is formed from two long polymer chains that are twisted around a common axis. The basic subunits are four specific nucleotides cytosine, guanine, adenine and thymine, which are ordered in a defined way. Nucleotides from both strands bind to form Watson-Crick base pairs. The connections between the two polymer chains are formed by hydrogen bonds between adenine-thymine and cytosine-guanine pairs. Adjacent base pairs are 3.4 Å apart and produce a rotation of 36°, so rather than forming a straight ladder, the double-helix structure is generated. The diameter of a DNA molecule is about 2 nm. One whole revolution is completed after 10 base pairs, giving a pitch of 3.4 nm. The whole repeat is formed from a major groove (2.2 nm) and minor groove (1.2 nm). The general structure of a double-helix DNA structure is shown in Figure 1, and AFM scans using the JPK NanoWizard® ULTRA Speed are shown in Figure 2 (see also [1]).
DNA forms many more complex structures in vivo and deformation of the “relaxed” B-DNA structure through bending or twisting is critical to the function and cooperation of many DNA-binding proteins. There are naturally occurring DNA motifs that extend the DNA self-assembly into a second dimension, for example the DNA replication fork is a three-way junction motif, and the Holliday junction is an example for a four-way junction motif. These more complicated structures suggested that DNA could be a promising candidate for nanotechnology.
DNA nanotechnology deals with the idea of designing and producing nucleic acid structures in a “bottom-up” approach. DNA is an excellent nano-construction material because of its inherent merits. The DNA sequence determines the hybridization, giving very predictable structures. It is fully biocompatible, can be copied and has the ability to be modified for specific applications; for example for nanoelectronics or biosensing. DNA self-assembles readily, which can be defined as a spontaneous association of components into organized structures using non-covalent interactions. DNA can be made into nanomachines, such as rotaxanes [2], or used to create tiling structures, such as the example in Figure 3.
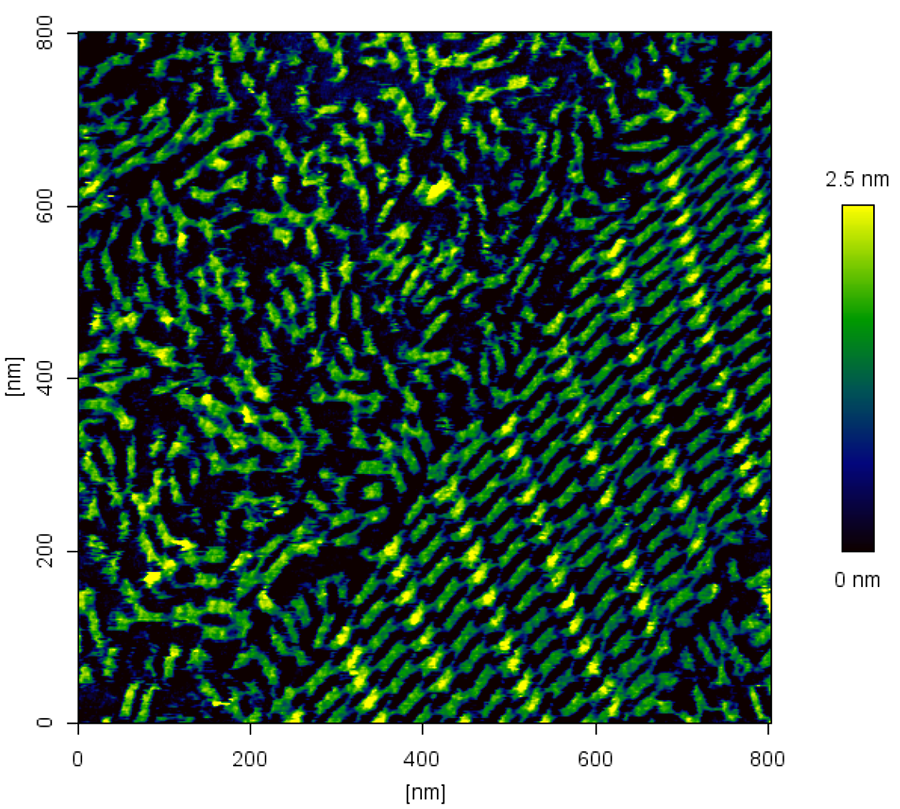
Tile-based DNA arrays
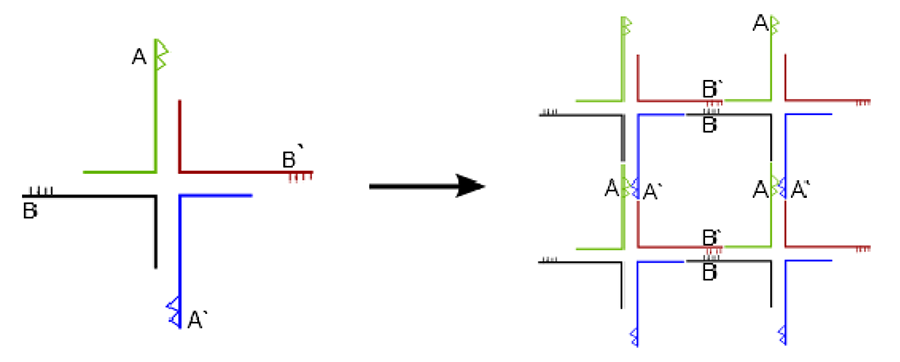
Seeman (see review, [3]) pioneered the concept of using DNA for nanotechnology by making complex DNA arrays. He proposed DNA “junctions” or “tiles” with complementary overhangs (sticky ends) which can self-assemble to create two-dimensional arrays. In the simplest form a four-arm junction can be created using four oligonucleotides with complementary sticky ends to create a periodic 2D array by self-assembly, as explained in Figure 4.
The size of the 2D array is in theory unlimited and depends only on the number of tiles. In really it was seen that formation of large arrays by simple junctions failed due to the inherent flexibility of the junction region. To solve this problem a large variety of complex DNA tiles with better stability have been developed and constructed.
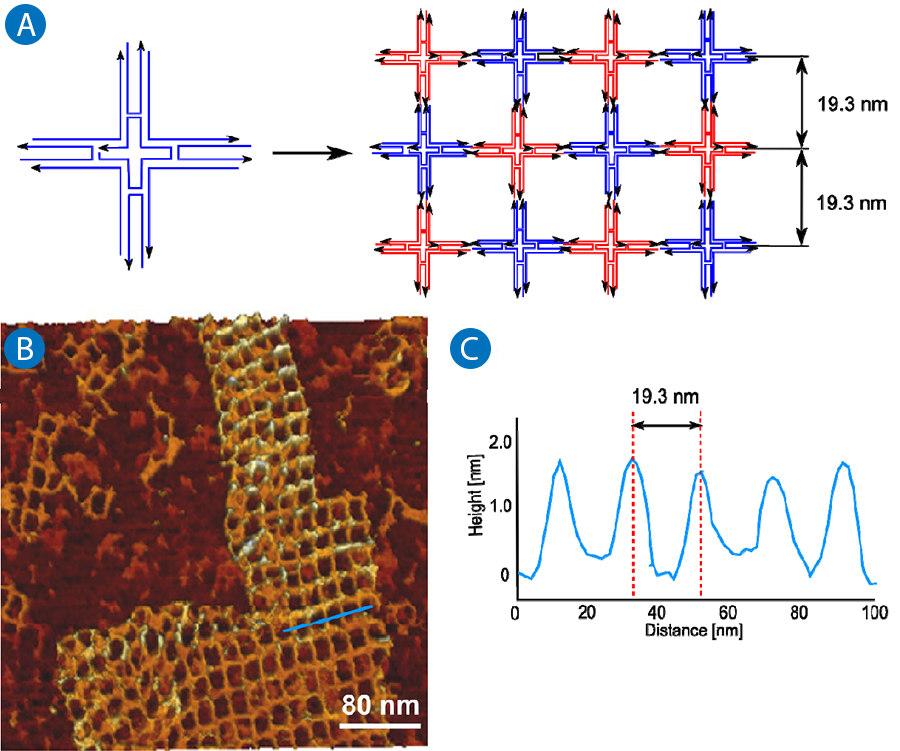
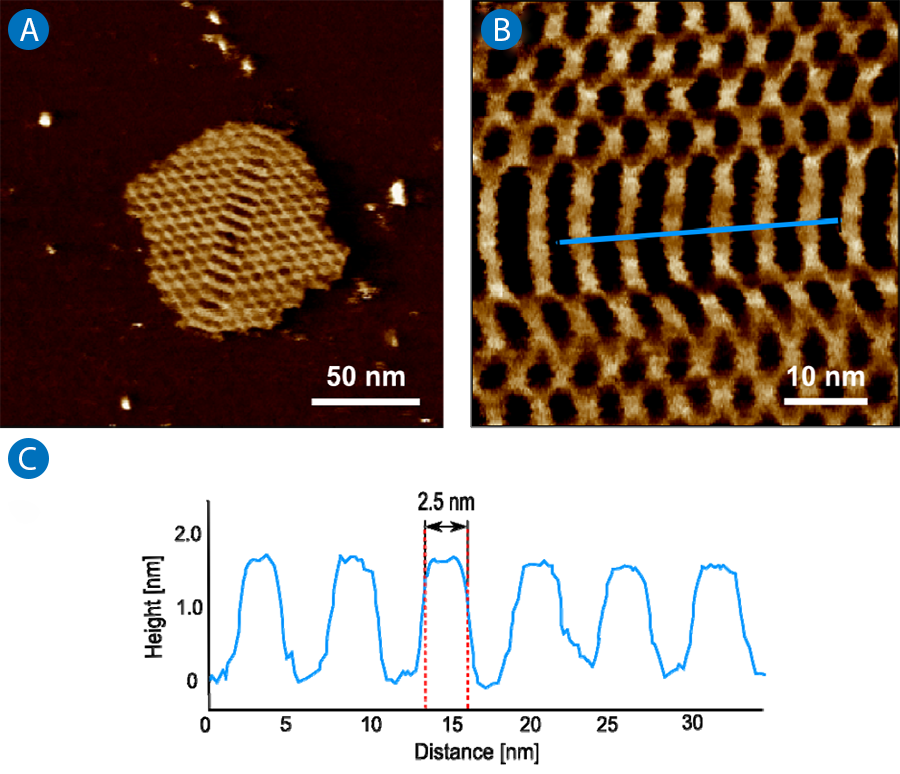
The 4 x 4 tile structure is one general example for a tile-based DNA array. The schematic explanation of the array is shown in figure 5A. The single tile consists of nine oligonucleotides, with one strand in the core of the tile which participates in all four junctions. The DNA junctions point in four directions with a 90° angle between every junction. The more complex arrangement within the building blocks provides more stability and the creation of larger 2D arrays.
The array is formed by alternating face up and face down tiles (shown in Figure 5A as blue and red respectively). The spacing within the 2D lattice is defined by the tile structures and should be 19.3 nm. Figure 5B and 5C show the results of the AFM measurements. One representative lattice can be seen in figure 5B. Next to large 2D structures also single and multiple tiles structures can be imaged that have not successfully self-assembled to lattices. The cross section in figure 5C demonstrates the expected spacing of 19.3 nm.
The sample was measured in TAE-Mg buffer on a mica surface without any fixation. The expected square grid structure with can be visualized in a very good quality without damage or distortion. To obtain a good image quality, the structure must be sensitively measured without damage. This is possible by using the JPK NanoWizard®3 or NanoWizard® ULTRA Speed, which can image with a very low tip-sample interaction using cantilever amplitudes below 2 nm.
Folding-based DNA origami
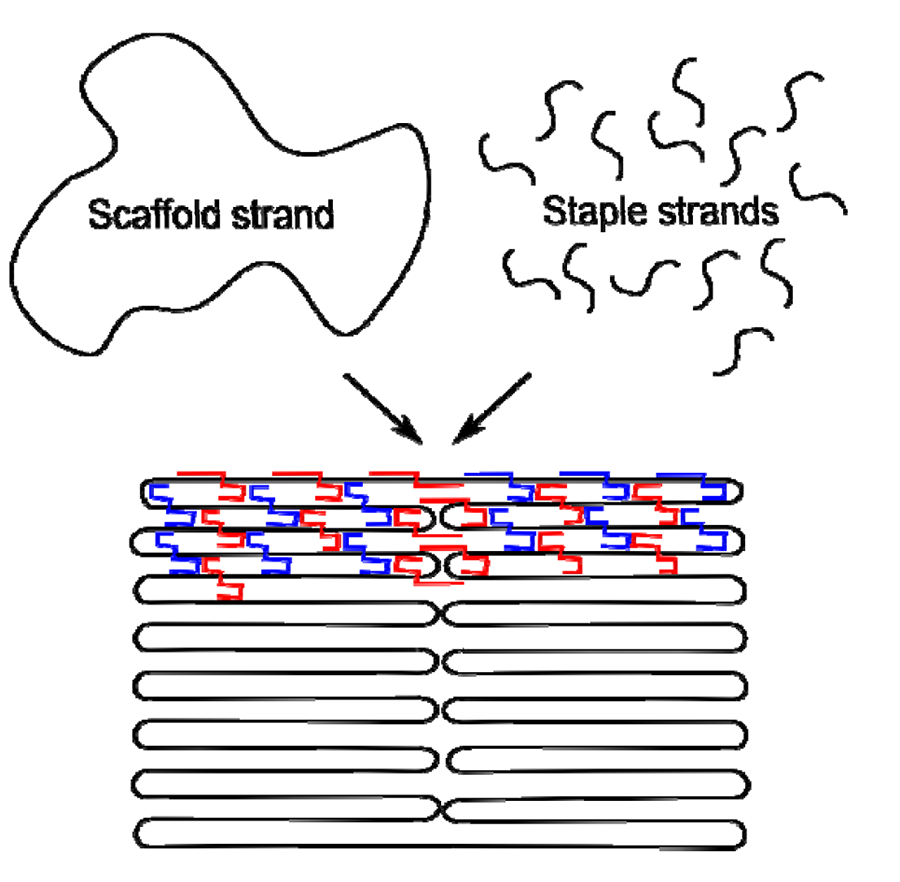
In general all the DNA nanostructures that are based on tiles belong to a multi-stranded approach. The structure is composed of short oligonucleotides with similar lengths. The idea of DNA origami, proposed by Rothemund [4], is to use one much longer DNA strand to provide a scaffold, which is folded within the structure and held by many complementary oligonucleotide strands. Rothemund used single-stranded viral DNA as the backbone for these structures, which were formed by folding it back and forth in a raster pattern to fill the desired shape. The shorter oligonucleotide “staples” hold the structure together forming antiparallel crossovers between adjacent raster lines of the backbone.
Figure 6 shows a schematic diagram explaining the process for a simple rectangle; origami patterns with other shapes can be produced with the same method. There are several advantages to having one long strand running through the whole structure. One obvious motivation is to improve the stability of the final structure. Even more importantly, the single continuous scaffold strand constrains the range of structures that could self-assemble and therefore helps the correct structures form more reliably, increasing the yield of properly-folded structures. AFM images of structures formed using this method are shown in Figure 7.
Another main advantage is the addressability of the locations within the structure. As can be seen in Figure 7B and C, the woven DNA strands form a lattice with around 5-6 nm spacing and can provide hundreds of uniquely defined positions for labelling. Since the final location of each staple strand is known within the origami structure, it is possible to add many different kinds of molecules, nanoparticles etc. to any position within the shape. Modified staples are used, which offer functional binding sites for other molecules or form shapes outside the structure in addition to the structural binding to the scaffold.
Rothemund [4] demonstrated the potential of the method by marking locations with DNA dumbbell hairpin staples. These staples had extra base pairs that bound within the same oligonucleotide rather than to part of the scaffold strand, forming a knot visible in AFM images. Further work from many groups has focused on labelling the structures with non-DNA components, for instance for fluorescence assays, or to form electrically active structures. Figure 8 shows rectangular origami structures before and after labelling with streptavidin.


The methods described here can be extended in various directions, depending on the particular focus within nanotechnology. For instance plasmonic structures can be created using biotinlylated DNA staples to bind streptavidin-coated metal nanoparticles. If the origami pieces are constructed with free sticky ends, they can tile into larger arrays of metal nanoparticles. For a more biomedical focus, multiple functional protein molecules can be bound separately or together in a defined pattern [5]. Alternative directions include making 3-dimensional shapes, as shown for instance in Figure 9.
The JPK NanoWizard® ULTRA Speed AFM also enables fast scanning of even delicate samples, as shown in Figure 10. As structures are created that have a stronger focus on dynamics, binding and structural changes in response to specific stimuli, it will be more important to image quickly to follow the dynamics.

References
- “Atomic force microscopy with nanoscale cantilevers resolves different structural conformations of the DNA double helix” C. Leung, A. Bestembayeva, R. Thorogate, J. Stinson, A. Pyne, C. Marcovich, J. Yang, U. Drechsler, M. Despont, T. Jankowski, M. Tschöpe, and B.W. Hoogenboom Nano Lett., 12: 3846-3850 (2012)
- “Design strategy for DNA rotaxanes with a mechanically reinforced PX100 axle” D. Ackermann, S.-S. Jester and M. Famulok, Angew. Chem. Int. Ed. 51: 1-6 (2012)
- “DNA NANOTECHNOLOGY: Novel DNA Constructions” N.C. Seeman, Annu. Rev. Biophys. Biomol. Struct. 27:225–248 (1998)
- “Folding DNA to create nanoscale shapes and patterns” P.W.K.Rothemund, Nature 440(16): 297-302 (2006)
- “Orthogonal Protein Decoration of DNA Origami” B. Saccà, R. Meyer, M. Erkelenz, K. Kiko, A. Arndt, H. Schroeder, K.S. Rabe and C.M. Niemeyer, Angew. Chem. Int. Ed. 49(49): 9378–9383 (2010)