Application Note: Optical Tweezing of Cells with the NanoTracker 2: Manipulation and Force Measurements
This application note highlights how the NanoTracker™ 2 optical tweezers system enables precise manipulation and force measurement of cells and particles. With piconewton sensitivity, multi-trap control, and integration with microfluidics, the system is ideal for studying cell mechanics, adhesion, signaling, and intracellular processes in real time.
More detailed information on the tapping mechanism itself and the working principle of optical traps can be found in our technical notes (e.g. "Quantitative force measurements with optical tweezers: The JPK NanoTracker™")
Contents include:
- Force measurements for adhesion, signaling, and membrane mechanics
- Intracellular transport, tether pulling, and viscoelasticity analysis
- Integration with microfluidics and fluorescence microscopy for live-cell studies

Introduction
In the last decades, optical traps (also known as optical tweezers) employing highly focused laser beams to trap and manipulate objects on the nano- and microscale have proven to be versatile tools in many biological and biophysical studies. The technique relies on the transfer of momentum from light entering a refractive particle which allows its stabilization in a trapping potential. From particles in the tens of nanometers range up to whole cells, a variety of objects can be held and accurately positioned. With advanced setups like JPK's NanoTracker™ 2, forces acting on the trapped objects can be measured with high precision down to single piconewtons and below. This opens up a broad range of possible experiments in the context of cellular research. Some fundamental applications will be introduced in this note.
More detailed information on the trapping mechanism itself and the working principle of optical traps can be found in our technical notes (e.g. "Quantitative force measurements with optical tweezers: The JPK NanoTracker™").

Cell/Bead manipulation
Since suspended cells typically have a higher refractive index than the surrounding medium, they can be trapped easily with optical tweezers. JPK's NanoTracker™ 2 offers various options for the independent positioning of multiple traps allowing the simultaneous manipulation of tens of particles or cells. The near-infrared laser source (wavelength 1064nm) ensures moderate absorption and heating of biological matter [1] which renders it a noninvasive tool perfectly suited for holding and positioning suspended cells in three-dimensional environments. Figure 1A shows yeast cells being gathered in a glass micro-capillary with NanoTracker™ 2 optical tweezers. With its powerful control software, the sorting of cells or their precise deposition on patterned growth substrates becomes as easy as "drag 'n' drop". The interaction of cells with functionalized surfaces is of great interest for research on biocompatible materials, e.g. for implants and antifouling surfaces in medical and technical applications. With the cell handling capabilities of the NanoTracker™ 2 system, cells, bacteria or other microorganisms can be selectively brought in contact with pretreated substrates to qualitatively evaluate their adhesion and growth behavior. Figure 1B shows a yeast cell that is optically trapped and approached to a vertical surface. The cell representing any type of micro-organism can be held in contact with the substrate in order to monitor its behavior in terms of adhesion or viability (in the case of anti-fouling treatments).
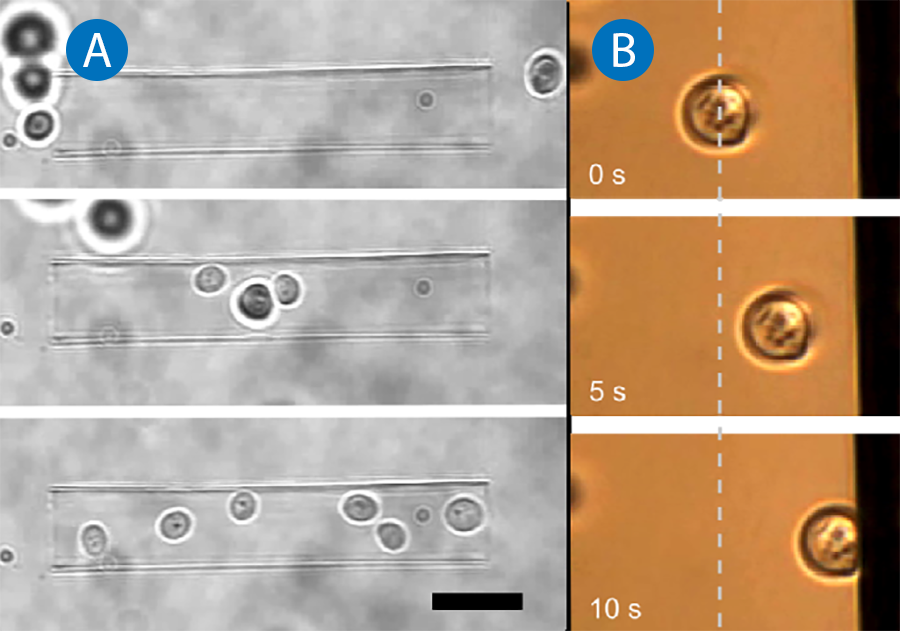
The range of possible measurements is further extended with the integrated microfluidic system of JPK's LaminarFlowCell (LFC™): stable parallel streams of different media containing different drugs or nutrients can be established. With the setup, cells can be exposed to these media and their response (e.g. in terms of altered mechanical properties as described below) can be monitored.
For simple cells like yeast or certain types of unicellular organisms that have well-defined shapes and optical properties, it is possible to calibrate the trap, i.e. to determine the trap stiffness and to measure forces acting on the organism. This is usually done by exposing the trapped cell to a known force and measuring its displacement from the stable trap position, e.g. by moving them through a viscous medium. This generates a drag force depending on the size and shape of the object as well as transport velocity, v, and medium viscosity, n. For a spherical cell or object (radius r) the force calculates as
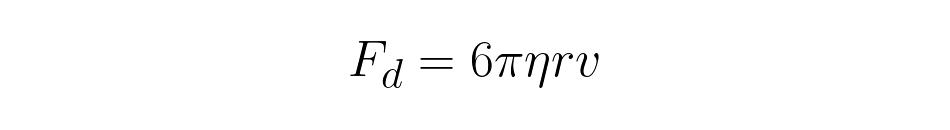
After successful calibration, external forces acting on the cell or binding forces in adhesion processes can be quantified and compared. For example, this was demonstrated for sickle-shaped Malaria parasites to measure adhesion forces during their surface binding activity [2].
Many complex cells are optically inhomogeneous or have irregular shapes even when in suspension. Thus the direct recording of absolute force values with such cells can be very demanding. A simple approach to overcome these inherent limitations is to use well-defined spherical particles, typically micrometer size polystyrene (PS) or silica beads attached to the cells as handles and force probes. Today, a large variety of beads differing in size, material, surface chemistry or fluorescent labeling is commercially available which enables researchers with little experience in surface modification to design and conduct complex experiments. This extremely useful toolbox and some of its applications will be introduced in the following sections.

Extra- and intracellular particle-cell interactions
The manipulation of cells and (functionalized) beads can be used to trigger certain biochemical reactions or to quantify the forces acting during cellular processes.
Beads carrying signaling molecules, receptors or other functional groups on their surface have frequently found application in studies investigating the interplay of such molecules with cellular signaling pathways. A basic experiment would be the triggering of cellular processes simply by bringing functionalized beads in contact with a cell. This has successfully been used to selectively activate single immune cells by presenting antigens to different regions of their surface [3].
Besides higher positioning accuracy compared to cells, the main benefit of using homogenous spherical beads in the trap is the ability to perform a precise calibration for high resolution displacement and force measurements. Briefly, with the NanoTracker™ 2, the Brownian motion of the particle in the trap potential is recorded and from the power spectrum of the fluctuations, the position sensitivity and trap stiffness are automatically calculated. For small displacements, the trap can be approximated as an elastic spring that exerts a force on the trapped object which is proportional to the object's displacement from the trap center. With this high precision force measurement tool at hand, numerous experimental configurations can be implemented.
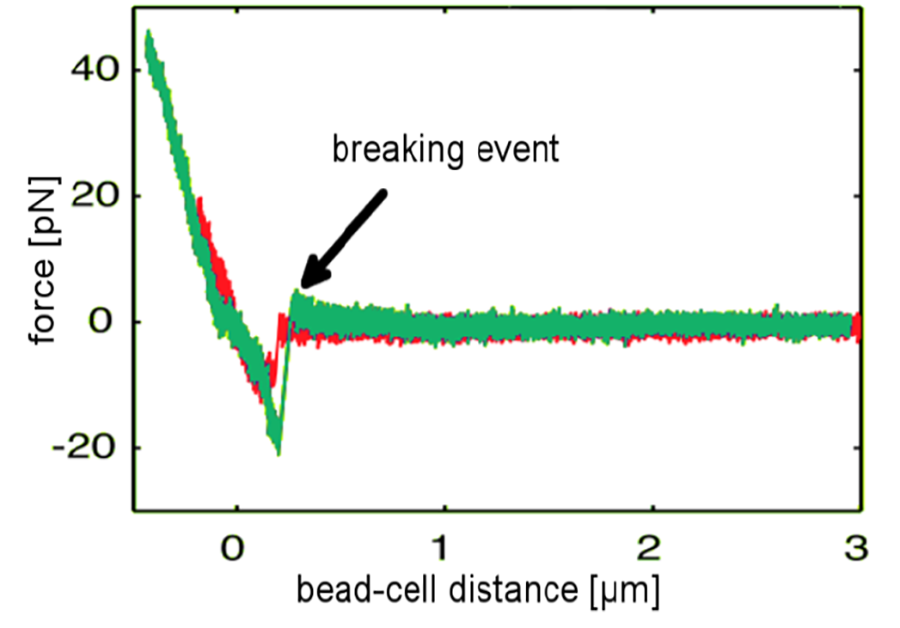
Calibrated systems with functionalized beads are used to investigate single binding kinetics of receptor-ligand, antigen-antibody or other binding partners on the surface of a cell. A functionalized bead is trapped and approached to the cell surface. After a defined contact time allowing the stochastic binding event to take place, the bead is pulled away from the cell and a forcedistance curve is recorded. Among other assays, this method was used to characterize the binding forces and dynamics of viral surface proteins to host cell receptors which play a decisive role in the development of infectious diseases and immune biology. Force-distance data recorded at different loading rates applied to the bond provides insights in the underlying binding mechanism. Figure 2 shows two representative curves (red and green) recorded with a NanoTracker™ setup as an influenza-virus coated bead is being pulled away from the cell surface. The characteristic jumps in the force signal correspond to individual bonds breaking apart [4].

It is important to note that force measurements and the manipulation of particles are not restricted to the cell surface. After a cell has engulfed a particle through endocytosis or a similar cellular uptake process, it can still be trapped and actively moved. Figure 3 shows a polystyrene bead being optically transported inside a macrophage cell. Intracellular force measurements require more elaborate calibration methods since the refractive index and viscosity of the cytosol surrounding the trapped particle need to be measured and taken into account. The inhomogeneity of the cell interior may contribute to larger errors in this type of measurement as well. Instead of particles, it is also possible to trap cellular organelles or other substructures directly. It was shown that the forces of kinesin motor proteins transporting lipid droplets along microtubules inside cells can be measured directly inside the living cell with optical traps after proper trap calibration [5]. Also rheological measurements determining the viscoelastic properties in different regions of the cell interior have been successfully performed with optical tweezers. Micro-scale granules in the cell can be trapped and oscillated at defined amplitudes and frequencies. From the displacement of the granules in the trap, detailed information on the loss and storage module (corresponding to viscous and elastic contributions) of the intracellular material can be derived [6-8].
Membrane properties
The outer cell membrane is a highly complex structure consisting mainly of a double layer of lipid and sterol molecules with embedded isolated or clustered proteins. It fulfills a variety of functions and directly interacts with other cellular components like the cytoskeleton. The mechanical properties of the cell membrane (tension, viscosity) are essential for many processes involved in cell signaling, motility or growth and abnormal changes are related to malfunctions and diseases. Thus, the characterization of the cell membrane under different environmental conditions or at different stages of cell proliferation and differentiation are of great interest for various research disciplines.
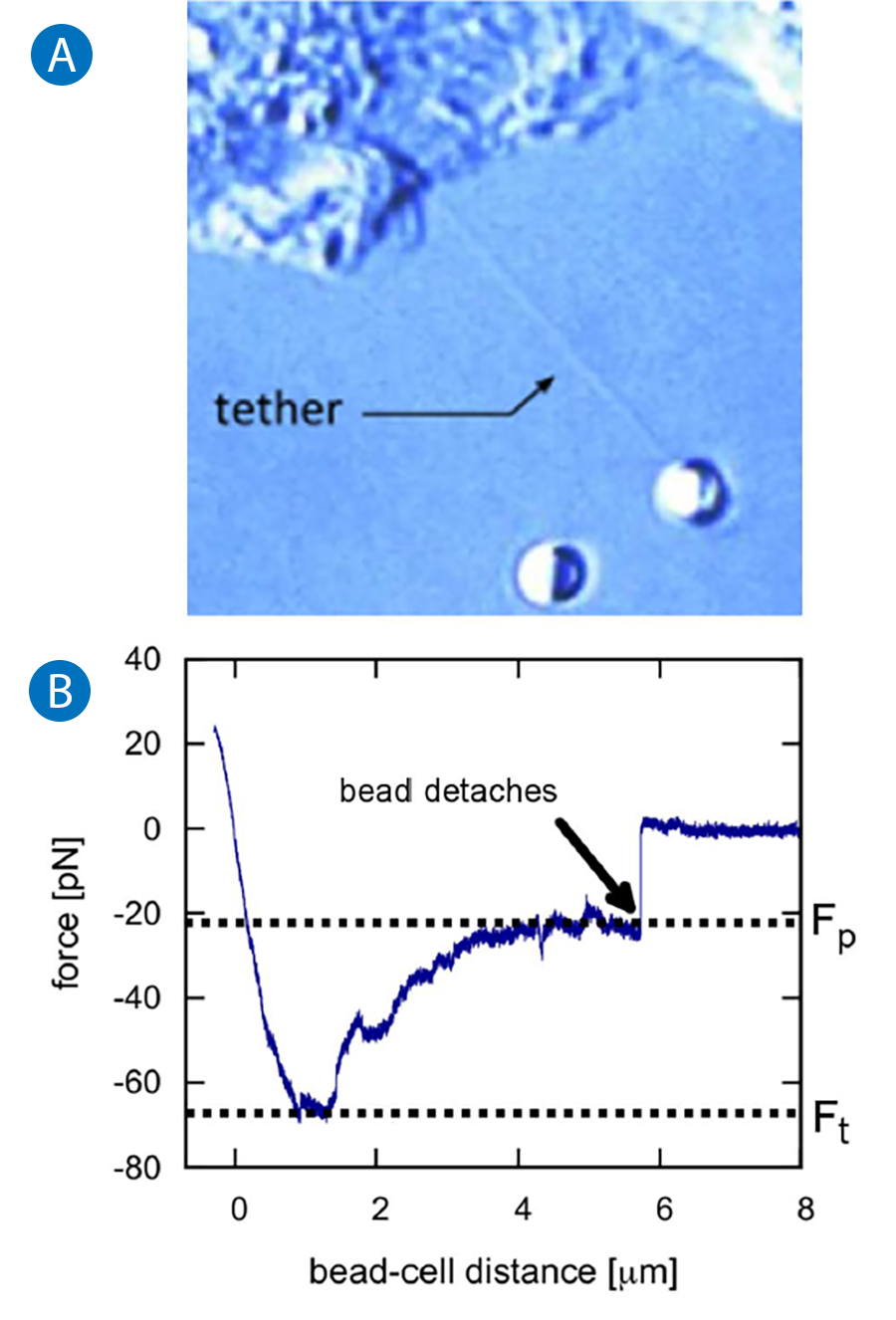
Information on local mechanical properties of the membrane can be derived from so called tether-pulling experiments. Beads coated with immunoglobulin G (IgG) antibodies are trapped and attached to the cell membrane. By applying force to the bead, several micrometer long membrane tubes (tethers) are pulled out while the forces on the attached particle are recorded (see figure 4 A). Typically, the so called tether formation force has to be exceeded before the tether forms. This threshold can be ascribed to membrane-cytoskeleton interactions that need to be overcome. Then a lower force plateau is reached where the tether is elongated with constant force since membrane reservoirs on the cell surface provide material for the growing tube.
The typical force-distance curve of a tether pulling process is displayed in Figure 4 B.
The plateau force, Fp , depends on the tether elongation speed and increases for higher velocities. Knowing Fp and the pulling velocity, v, allows the calculation of an effective membrane viscosity neff [9]:
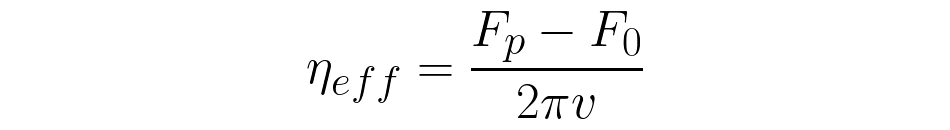
and effective membrane tension yeff [10]:
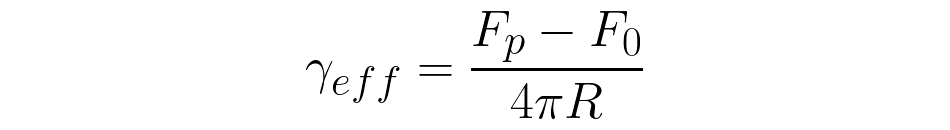
where F0 is the force required to hold the tether stable at zero velocity (v=0) and R is the thether's cross-sectional radius.
Results from tether formation experiments have provided detailed insight in mechanisms as diverse as cell motility, vesicle trafficking and intercellular signaling and are currently in the focus of many research labs [11].
Cell mechanics
Numerous studies have found evidence for a close relationship between the mechanical properties of cells and their function. The most prominent examples are the malignancy and invasiveness of cancer cells or different erythrocyte related diseases that are directly linked to the mechanical stiffness and viscoelasticity of the respective cells [12-14].
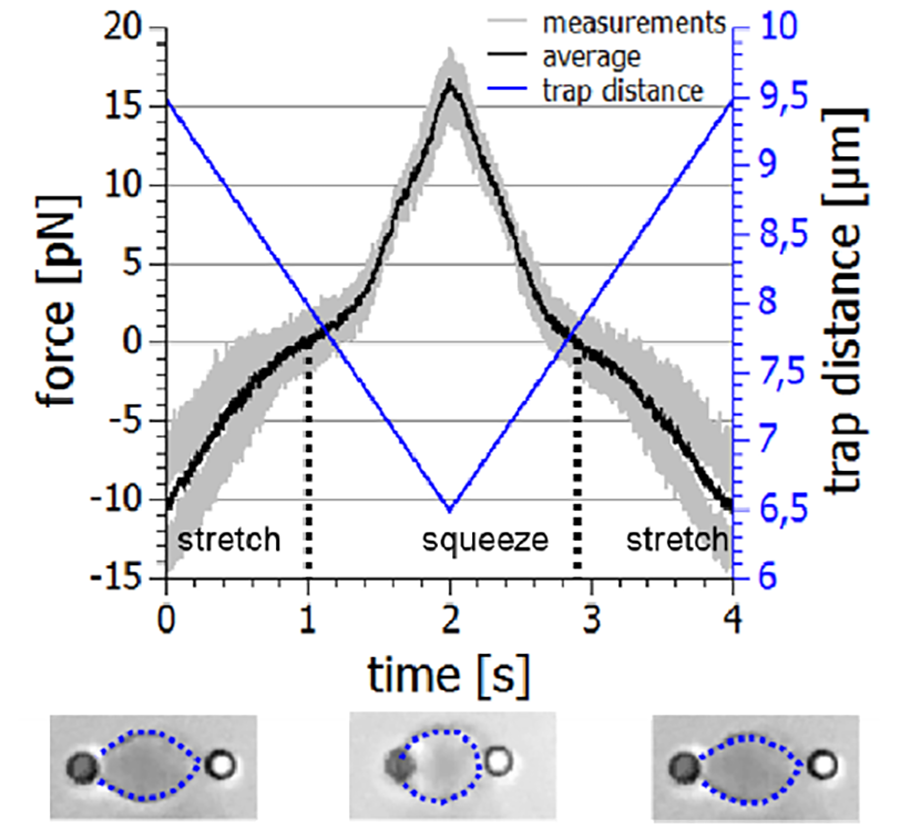
Single cell rheology is thus increasingly recognized as a powerful tool to identify different cell types or to detect cellular (mal-)functions. While the mechanical properties of cells organized in tissue or strongly adhered to rigid substrates can be measured with bulk rheology or atomic force microscopy (AFM) methods, these techniques are not suitable for freely floating cells that reside in the blood or lymph. Their native environment is a fluid they are suspended in which makes it difficult to evaluate their viscoelasticity under physiological conditions. JPK's NanoTracker™ 2 provides an excellent alternative to investigate the mechanics of suspended cells under welldefined experimental conditions. The cells can be decorated with appropriately functionalized beads that bind to the cell surface or the underlying cytoskeleton, depending on the focus of interest. Two beads located diametrically opposite on the cell surface provide ideal handles for optical tweezers to stretch or squeeze the cell with adjustable frequencies and amplitudes, and to record the viscoelastic response of the system.
Figure 5 shows a typical example of an erythrocyte mechanics experiment. The cell is held with two beads in a dual optical trap. By moving one of the traps, in this case with acousto-optical deflectors (AODs), the cell is repeatedly stretched and squeezed while the forces acting on the beads are recorded. From the forcedistance relation plotted in figure 5, important information on the mechanical properties of the cell and thus its physiological state can be derived. In addition to live blood cells, so called erythrocyte ghosts consisting mainly of the cell's membrane can be investigated in order to evaluate the effect of different drug treatments on membrane properties [15,16].
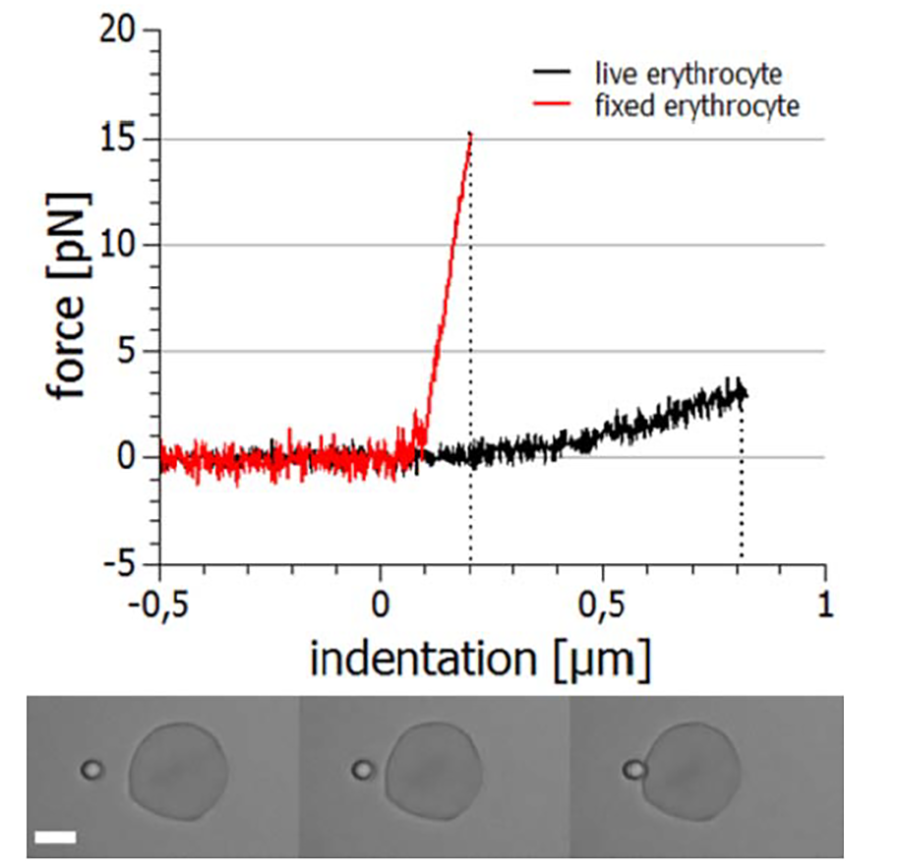
Other aspects of cellular viscoelasticity can be investigated with experiments where a cell that is fixed in space (e.g. aspirated by a micropipette or adhered to a surface) is indented with a microparticle [13]. The indentation depth is typically in the range of tens to hundreds of nanometers which requires highly stable and accurate positioning and force measurement instrumentation as it is provided by the NanoTracker™ 2 system. In figure 6 the course of such an experiment and the recorded forces of the approach and retraction segments are displayed. It is obvious that indenting a live cell requires much lower forces (approx. 3 pN for 800 nm) than indenting a fixed cell (approx. 15 pN for 200 nm) which is due to the stiffness increase caused by the fixation agent.
These and similar experiments deliver important insight into the link between biological function and mechanical properties of cells. This helps to identify potential targets for novel drugs that regulate cell function by influencing cell mechanics and the related dynamics.
Conclusion
Versatile, highly developed optical tweezers and detection systems like JPK's NanoTracker™ 2 can be used for a broad range of cell-related experiments from cell sorting and single-molecule interactions to the investigation of whole-cell mechanics. The complex effects that drugs and functional molecules have on cellular behavior and mechanics can be studied with this advanced system, and the integration of microfluidics even extends the range of addressable questions. New applications, especially in combination with high-resolution fluorescence microscopy, are continuously developed to shed more light on the fascinating interplay between cellular mechanics, dynamics, and signaling. For further reading on these topics, we recommend our technical notes on fluorescence and confocal integration with the NanoTracker™ optical trapping system.
Literature
- Y. Liu, D. K. Cheng, G. J. Sonek, M. W. Berns, C. F. Chapman, and B. J. Tromberg, Biophys. J. 68, 2137 (1995).
- S. Hegge, K. Uhrig, M. Streichfuss, G. Kynast-Wolf, K. Matuschewski, J. P. Spatz, and F. Frischknecht, ACS Nano 6, 4648 (2012).
- X. Wei, B. J. Tromberg, and M. D. Cahalan, Proc. Natl. Acad. Sci. 96, 8471 (1999).
- C. Sieben, C. Kappel, R. Zhu, A. Wozniak, C. Rankl, P. Hinterdorfer, H. Grubmuller, and A. Herrmann, Proc. Natl. Acad. Sci. 109, 13626 (2012).
- G. T. Shubeita, S. L. Tran, J. Xu, M. Vershinin, S. Cermelli, S. L. Cotton, M. A. Welte, and S. P. Gross, Cell 135, 1098 (2008).
- M. Yanai, J. P. Butler, T. Suzuki, A. Kanda, M. Kurachi, H. Tashiro, and H. Sasaki, Am. J. Physiol. - Cell Physiol. 277, C432 (1999).
- C. Selhuber-Unkel, P. Yde, K. Berg-Sørensen, and L. B. Oddershede, Phys. Biol. 6, 025015 (2009).
- L. G. Wilson and W. C. K. Poon, Phys. Chem. Chem. Phys. 13, 10617 (2011).
- F. M. Hochmuth, J. Y. Shao, J. Dai, and M. P. Sheetz, Biophys. J. 70, 358 (1996).
- I. Titushkin and M. Cho, Biophys. J. 90, 2582 (2006).
- A. Diz-Muñoz, D. A. Fletcher, and O. D. Weiner, Trends Cell Biol. 23, 47 (2013).
- T. W. Remmerbach, F. Wottawah, J. Dietrich, B. Lincoln, C. Wittekind, and J. Guck, Cancer Res. 69, 1728 (2009).
- Z. L. Zhou, B. Tang, and A. H. W. Ngan, Nano LIFE 02, 1250010 (2012).
- G. A. Barabino, M. O. Platt, and D. K. Kaul, Annu. Rev. Biomed. Eng. 12, 345 (2010).
- J. Sleep, D. Wilson, R. Simmons, and W. Gratzer, Biophys. J. 77, 3085 (1999).
- M. Dao, C. T. Lim, and S. Suresh, J. Mech. Phys. Solids 51, 2259 (2003).

