Novel EPR Hybrid Technique Provides Greater Detail on the Structure of Membrane Proteins
EPR analysis demonstrated the tight packing of the two YgaP second transmembrane helices upon binding of the catalytic product
Membrane proteins are critical to cell survival, enabling material transportation, signal transduction, and energy metabolism across the cell membrane. The functions of such proteins have been largely elucidated through detailed studies of their structure. High resolution images of protein structure can be obtained using X-ray diffraction and cryo-electron microscopy. In order to observe changes in protein structure in response to binding, nuclear magnetic resonance (NMR) is typically needed. Unfortunately, NMR is limited to use on relatively small protein structures. Such size limitations have precluded determination of the tertiary fold of full-length YgaP.
YgaP is an E. coli integral membrane sulfurtransferase. It belongs to a large superfamily of enzymes found in bacterial, archaeal, and eukaryotic cells. Such enzymes are responsible for the transfer of sulfur between the cell and its environment. Sulphur is essential for a range of functions that support life, and enzymes involved in sulfur transfer reactions are critical for maintaining homeostasis.
The three-dimensional topological structure of a full-length E. coli dimeric YgaP has now been described using electron paramagnetic resonance (EPR) in combination with site-directed spin labelling (SDSL) and computational approaches. EPR has a much greater sensitivity than NMR and is therefore not limited by the size of a protein. Continuous-wave-EPR spectra were acquired using a Bruker A300 spectrometer (Bruker Biospin GmbH, Rheinstetten, Germany) equipped with a high-sensitivity cavity (ER 4119HS, Bruker Biospin GmbH, Rheinstetten, Germany). Double electron-electron resonance (DEER) experiments were performed with a Bruker Elexsys E680 spectrometer (Bruker Biospin, Billerica, MA).
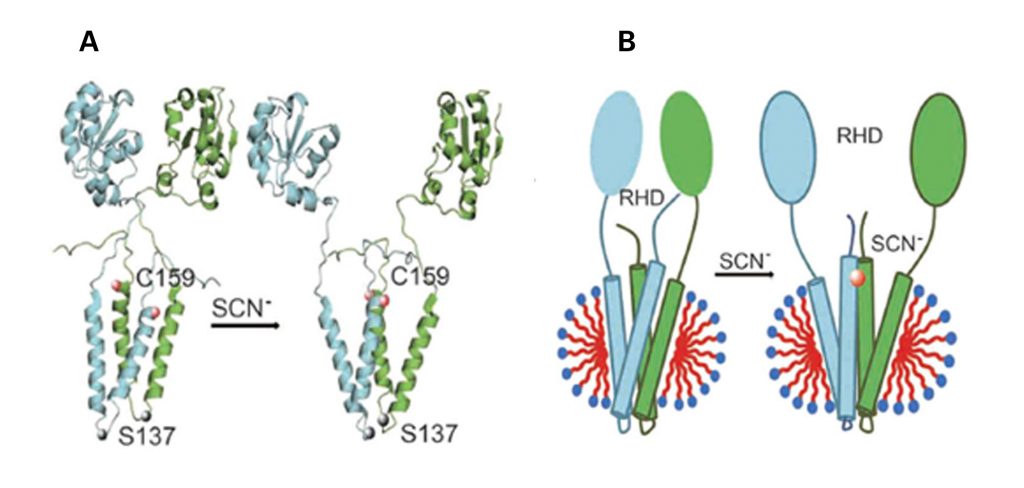
The previous structural studies of YgaP showed that YgaP monomers comprise a cytosolic catalytic rhodanese domain and a transmembrane domain (TMD). Now mobility, accessibility and membrane immersion measurements, made through EPR analysis, have revealed a helix-loop-helix secondary structure of the YagP-TMD monomers. Furthermore, measurements both within and between the monomers in conjunction with rigid-body computation have determined the structure of the tertiary folds of both the YgaP-TMD and full-length YgaP. Additional EPR analysis of the arrangement of the transmembrane helices after binding of the catalytic product, thiocyanate, provide insight into the dynamic properties of YgaP in the E. coli membrane.
These new structural details suggest that YgaP plays a crucial role in thiocyanate exportation from E-coli cells and may undergo conformational rearrangement and mobility changes on binding thiocyanate. Further functional analysis and spectroscopic experiments will be needed to fully describe the YgaP exportation mechanism.
Figure 1. YgaP structure before and after binding thiocyanate: (a) Hypothetical conformational changes (b) Topological model in DPC micelles
RHD=rhodanese domain; SCN= thiocyanate
Ling, S. et al. Structure of an E. coli integral membrane sulfurtransferase and its structural transition upon SCN− binding defined by EPR-based hybrid method. Sci. Rep. 6, 20025; doi: 10.1038/srep20025 (2016). © Creative Commons Attribution 4.0 International License http://creativecommons.org/licenses/by/4.0/