Neuroscience Research
Chemical synapses in the human brain are small
Due to properties like size and molecular density, synapses and other structures of interest to neuroscience researchers have been difficult to study by conventional methods of microscopic investigation. Yet extracting information about the spatial organization, stoichiometry, interactions, and dynamics of these structures on a molecular level is essential; advancing our understanding of the human brain depends on developing more and better methods to do so.
Ideal neuroscience microscopy methods should not only achieve higher resolutions than conventional means, but also distinguish specific structures and provide complete and reliable information for multiple sample types.
In providing exactly these capabilities, super-resolution microscopy with single-molecule localization (SMLM) has overcome many of the limitations of conventional neuroscience microscopy. This has already enabled dramatic improvement of our understanding of neuronal and brain function and will continue to create new opportunities for investigation and discovery in the field going forward. Only the Vutara VXL provides unparalleled SMLM capabilities for neuroscientists.
TABLE OF CONTENTS
Are you interested in Bruker's best-in-class solutions for super-resolution SMLM?
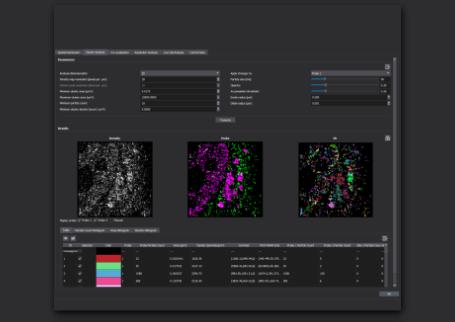
Image Gallery: SMLM Applications in the Neurosciences
Retinal Tissue Slice to Study Neural Development
MAGENTA: CALBINDIN; CYAN: PSD95

Retinal Tissue Slice to Study Neural Development
CALBINDIN
Retinal Tissue Slice to Study Neural Development
PSD95

C. elegans as Model System for Synaptic Transmission

C. elegans as Model System for Synaptic Transmission

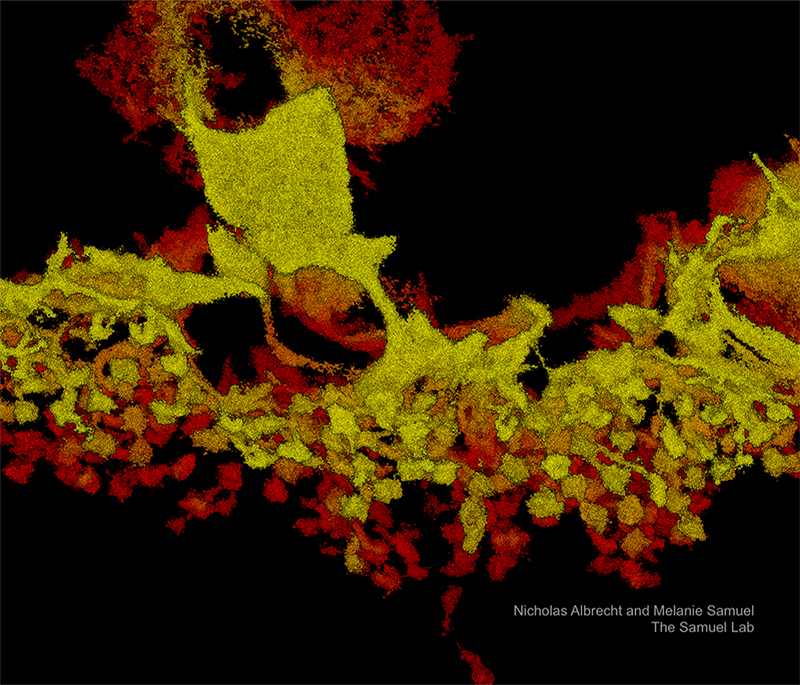
Retinal Synapses as imaging by the Vutara VXL

Axons from primary rat hippocampal neurons

SMLM vs. Other Neuroscience Microscopy Methods
Neuroscience researchers face a unique challenge when selecting an imaging methodology: historically, no single method has been able to provide complete information about neuronal structures and processes at the molecular level, so selecting one has required compromising between capabilities or conducting complementary investigations via multiple methods.
For example, many structures of interest are barely visible or undiscoverable by methods that are capable of specific labeling, like diffraction-limited two-photon microscopy. Yet electron microscopy, which provides superior resolution and is the only method capable of visualizing structures of interest smaller than 50 nm — like the synaptic cleft and synaptic vesicles — lacks fluorescence microscopes' capability for easy protein-labeling of specific structures.1,2 This restricts researchers' opportunities to perform complex investigations, including protein identification or neuronal tracing.
How is Super-Resolution SMLM Different?
Contrasting conventional methods of neuroscience microscopy, super-resolution microscopy both enables target-specific fluorescence labeling and achieves resolutions well below the diffraction limit of light microscopy. As a result, it can provide unique and often otherwise inaccessible insights into structures and processes.
Among the various super-resolution imaging techniques used in neuroscience research — including structured-illumination microscopy (SIM), stimulated emission-depletion (STED), and expansion microscopy — SMLM is the only method that relies entirely on analysis of the molecules imaged. SMLM also provides the highest resolution.3 As a result, with its lateral resolution of 20 nm and specific fluorescence labeling, SMLM is an exquisite tool to advance neuroscience.
Find out more on our 3D Imaging with Bi-Plane page.
Practical Considerations for Super-Resolution SMLM in Neuroscience Applications
What is SMLM Used for in Neuroscience Research?
As a result of its advantages in labeling specificity and resolution, SMLM has the potential to be highly applicable to many questions of interest to neuroscience researchers and SMLM techniques have already been used in a broad range of neuroscience applications.
To date, the results of these studies have demonstrated the applicability of the method to both structural and quantitative analysis as well as 3D imaging of both tissue and cultured samples.3 In these applications, SMLM has been able to provide insight into the spatial organization, stoichiometry, and temporal dynamics of a broad range of structures and processes and provides significant advantages over other methods for investigating molecular interactions (molecular size, density, distance apart, overlap, etc.).
Does SMLM Have Limitations for Neuroscience Research?
SMLM studies demonstrated on tissue samples appear less frequently in SMLM literature than those demonstrated on cultured cells. This imbalance is especially apparent for applications that call for 3D imaging in tissue, and can be attributed to several sources of frustration for researchers. These include:
- Sample preparation is more complex and time-consuming for tissue samples than for cultured cells
- Tissue samples demonstrate more — and more disruptive — background signals from fluorescent molecules3 (and would require more advanced analytical tools to process)
As a result, the achievable imaging depth of SMLM techniques can create challenges for working with tissue slices and thick samples.*
*Some instrumentation achieves superior imaging depth, especially for 3D imaging in tissue samples
(see "Advantages of Bruker's Vutara VXL for Neuroscience Applications" for more information).
SMLM is applicable to the study of neuronal structures and processes, including (among others):
- Neural development
- Neural circuit assembly
- Synaptic formation
- Synaptic transmission
- Synaptic proteins
Recommendations for SMLM-Capable Neuroscience Microscopes
What You Need to Know About SMLM for Neuroscience Applications
Depending on the equipment, sample, and fluorophore is used, SMLM methods have the potential to achieve high lateral localization precisions and ~20 nm spatial resolution. Yet the actual resolution and localization precision achieved when imaging both cultured samples and tissue slices depend on a variety of additional factors. These include, among others, labeling density and accuracy, signal size and strength, and microscope drift. Optimizing sample praparation methods for SMLM by, for example, increasing labeling antibody concentration, can resolve many of these issues and clear the way for successful analysis.
Why Tool Selection Is As Important As Technique
The components and functionality of an SMLM imaging system itself can limit the resolution and localization precision achievable by any of the various SMLM techniques — especially when performing 3D imaging in tissue slices. Moreover, SMLM-capable microscopes can require time-consuming setup that distracts from critical research processes. They also often pair with analytical tools that lag behind their imaging capabilities.
Ideal SMLM-Capable Neuroscience Microscope Features
Given the potential for instrument configuration to arbitrarily constrain the achievable results of SMLM investigation, it is important to select equipment with known capabilities that align with your specific research goals. Features frequently credited with providing the greatest opportunity for success and enabling researchers to take full advantage of the benefits of SMLM for neuroscience applications include:
- Dense and specific fluorescence labeling
- Multiplexing capabilities
- Enhanced imaging depth
- Advanced analytical support
Advantages of Bruker's Vutara VXL SMLM Microscope for Neuroscience Research:
Bruker's Vutara VXL offers best-in-class ease of use and imaging depth, supporting a broad range of different experimental needs. Key features include:
- Automated Workflows:
SRX software makes it easy to acquire the necessary data sets. Workflows allow the user to focus on the biological question instead of tinkering with complicated microscope settings. - Superior 3D Imaging & Sample Flexibility:
Proprietary bi-plane detection provides three-dimensional information and enables imaging within tissue slices. - Unlimited Multiplexing Capabilities:
The integrated Microfluidics Unit allows sequential labeling of an unlimited number of fluorescent probes. - Expert Applications Support:
Vutara VXL users can receive personalized guidance for sample prep optimization specific to their research objectives.